Auditory Event-Related Dynamics of the EEG Spectrum and Effects of Exposure to Tones
Scott MakeigDownload this reprint(.pdf, 38K).
Disclaimer: This work was supported by the Department of the Navy, Naval Medical Research and Development Command, Bethesda, Maryland. The views expressed in this article are those of the authors and do not reflect the official policy or position of the Department of the Navy, Department of Defense, nor the U.S. Government. Approved for release, distribution unlimited.
ABSTRACT
A new measure of event-related brain dynamics, the event-related spectral perturbation (ERSP), is introduced to study event-related dynamics of the EEG spectrum induced by, but not phase-locked to, the onset of the auditory stimuli. The ERSP reveals aspects of event-related brain dynamics not contained in the ERP average of the same response epochs. Twenty-eight subjects participated in daily auditory evoked response experiments during a four day study of the effects of 24-hour free-field exposure to intermittent trains of 89 dB low frequency tones. During evoked response testing, the same tones were presented through headphones in random order at 5 s intervals. No significant changes in behavioral thresholds occurred during or after free-field exposure. ERSPs induced by target pips presented in some inter-tone intervals were larger than, but shared common features with ERSPs induced by the tones, most prominently a ridge of augmented EEG amplitude from 11 to 19 Hz, peaking 1-1.5 s after stimulus onset. Following 3-11 hours of free-field exposure, this feature was significantly smaller in tone-induced ERSPs; target-induced ERSPs were not similarly affected. These results therefore document systematic effects of exposure to intermittent tones on EEG brain dynamics even in the absence of changes in auditory thresholds.
INTRODUCTION
Limitations of the Event-Related Potential (ERP)
Most studies of brain electrical activity recorded from the scalp are concerned either with analysis of differences in the mean spontaneous EEG frequency spectrum under various conditions, or with measures on averages of precisely timelocked brain responses evoked by discrete experimental stimuli or events. The low amplitude of most evoked-response components recorded from the scalp makes study of their parameters in single trials difficult or impossible. By averaging response epochs timelocked to a number of similar events, rhythmic or quasi-rhythmic brain activity not exactly synchronized in both time and phase to the experimental events is averaged out through phase cancellation, and the coherent time-and-phase-locked activity that remains is called an event-related potential (ERP). Since the development of computer averaging technology in the 1960's, the average evoked response method has become predominant for human experiments.
However, the electrophysiological response of the brain to auditory events is not fully captured by the ERP. This would only be the case if, (1) evoked responses were stable across trials and, (2) were completely independent of the ongoing EEG, and, (3) if the ongoing EEG were not itself perturbed by experimental events. Several reports have shown that ERPs are neither stable nor wholly independent of the EEG (e.g. Basar, 1980; Jansen and Brandt, 1991), and therefore, the response of the brain to auditory or other experimental events is not fully captured in the ERP.
Efforts to observe, quantify, and best characterize phase-incoherent event-related brain dynamics is an emerging research topic (Pfurtscheller, 1992; Van Dijk, 1992; Achimowicz, 1992). Violations of independence of the EEG and ERP can be modeled either as effects of experimental events on the EEG, and/or as non-stationarity in the ERP itself. That is, the difference between any single response epoch and the average ERP may be modeled either as being a perturbed continuation of pre-baseline EEG activity, as variance in the evoked response, or as some combination of these.
The Event-Related Spectral Perturbation (ERSP)
In modeling event-related reactivity in the ongoing EEG, amplitude and phase effects may be considered separately or in combination. A well-known example of an event-related EEG amplitude change is the reduction in occipital alpha-band EEG amplitude that normally occurs when the eyes are opened. The timecourse of event-related attenuations in alpha-band EEG was first quantified by Pfurtscheller (1977), using a term, event-related desynchronization (ERD), derived from the concept that cortical arousal involves "desynchronization" of relatively slow, spatiotemporally coherent cortical rhythms, and their replacement with faster, more spatially differentiated activity. ERD analysis was first used to quantify the degree to which the amplitude of prominent ongoing ("synchronized") alpha band EEG was depressed (or "desynchronized") following arousing events in an auditory contingent negative variation (CNV) paradigm, the time course and scalp topography of this depression depending on the stimuli and subject task and conditions (Pfurtscheller and Klimesch, 1991). Recently, Pfurtscheller (1992) has proposed the term event-related synchronization for timelocked amplitude enhancement, and Van Dijk et al. (1992) have analyzed background reactivity in various pre-defined frequency bands.
The measure introduced here, the event-related spectral perturbation (ERSP), can be viewed as a generalization of the ERD. The ERSP measures average dynamic changes in amplitude of the broad band EEG frequency spectrum as a function of time relative to an experimental event. That is, the ERSP measures the average time course of relative changes in the spontaneous EEG amplitude spectrum induced by a set of similar experimental events. These spectral changes typically involve more than one frequency or frequency band, so full-spectrum ERSP analysis yields more information on brain dynamics than the narrow-band ERD.
To compute an ERSP, baseline spectra are calculated from the EEG immediately preceding each event. The epoch is divided into brief, overlapping data windows, and a moving average of the amplitude spectra of these is created. Each of these spectral transforms of individual response epochs are then normalized by dividing by their respective mean baseline spectra. Normalized response transforms for many trials are then averaged to produce an average ERSP, plotted below as relative spectral log amplitude on a time-by-frequency plane.
The current study
Prolonged exposure to intense sound can produce temporary or permanent hearing loss and other physiological effects (Clark, 1991; Jansen, 1991), as well as emotional stress and cognitive performance decrements (Dejoy, 1984; Jones and Broadbent, 1991). Pathological effects of noise are often assumed to begin with temporary exposure-related increases in thresholds, termed temporary threshold shifts (TTS) (Melnick, 1991). These are accompanied by changes in auditory nerve activity and in auditory brainstem responses (ABRs) recorded from the scalp (Attias and Pratt, 1985). However, the effects of sound exposure on later evoked response features and on the EEG spectrum do not seem to have received much attention. This paper explores the effects of sound exposure on patterns of auditory event-related EEG spectral reactivity. It reports results of an experiment conducted during a larger study of the effects of exposure to active sonar signals (Coyne, 1992) investigating the effects on EEG spectral reactivity of 24 hours of intermittent free-field exposure to low frequency tones delivered at a level and duty cycle insufficient to induce permanent or temporary threshold shifts in any subject. Results demonstrate that characteristic event-related perturbations of EEG spectral amplitudes (ERSPs) are induced by both tones and tonepips. Prolonged exposure to tones are found to produce changes in this auditory spectral reactivity, even in the absence of behavioral threshold shifts.
METHODS
Subjects
Four groups of 10-12 normal hearing young adult volunteers (ages 19 to 34 years) were housed for four days in an enclosed laboratory setting. On each day, a fixed schedule of experiments was conducted, consisting of repeated audiograms, computerized cognitive tests, and physiological measures including the daily evoked response experiment reported here. Data from the 28 subjects who completed all portions of the experiment were used in the final analysis.
Exposure
Beginning at 8 a.m. of the third day, and continuing for 24 hours until 8 a.m. of the fourth day, trains of 6 consecutive 1 s pure tones were delivered in the free-field of the subjects' living quarters via multiple loudspeakers with an inter-train interval of 24 s. Tone frequencies were 720, 800, 880, 960, 1040 and 1120 Hz. Each tone had a linear 25 ms onset and offset. During exposure six different trains consisting of these 6 frequencies in different orders were presented continuously, at 30 s intervals, in cyclic order. Intensity of free-field stimulation was 89 dB (A-weighting). Free-field sound intensities were made uniform by careful balancing of multiple loudspeakers (Krebs et al., 1992). Subject dosages were verified by audio dosimeters worn by some of the subjects. Free-field exposure occurred only on Day 3, on Days 1, 2, and 4, no free-field stimulation was given.
Testing Procedure
Each of the four days of the session, subjects participated in a 15 minute experiment in which the same 1 s tones delivered in the free-field were delivered to the subject binaurally via headphones at 89 dB. Subjects were tested one at a time, in the same order each day, the experiments lasting from about 11 a.m. to 6 p.m. Each day at the time of testing, subjects would first be fitted with EEG electrodes. Then they would leave their free-field exposure living quarters and walk 50 feet to the evoked response testing chamber. Testing would begin within a few minutes after arrival.
Test Stimuli
During the experiment, three concurrent streams of auditory stimuli were delivered to subjects: the 1 s tones; occasional brief, high-frequency tone pips which served as target stimuli; and a continuous steady-state stimulus train at 40 dB normal hearing level (nHL). These are illustrated in Figure 1.
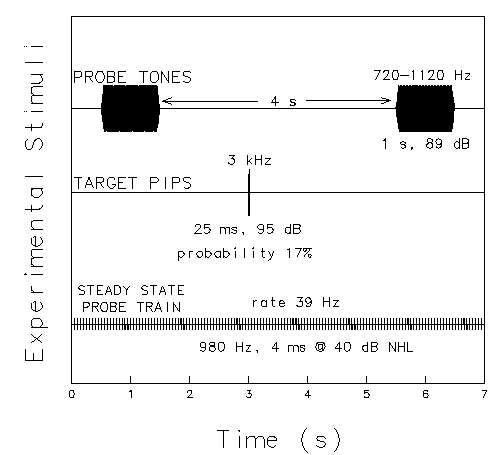
Figure 1. Schematic view of the three streams of test stimuli used in the experiment. The top trace shows 1s tones which are presented at regular 5 s intervals. Tones of six frequencies are delivered in pseudo-random order. In one sixth (17%) of the trials, a brief, high-frequency tone pip (middle trace) is presented at a random point during the inter-tone interval. Subjects press a response button whenever the target pip is presented. In the background (bottom trace), a continuous 39 Hz steady-state response probe train is presented throughout the experiment.
The tones were identical to the tones used in free-field exposure. However, because of technical limitations, instead of being delivered in trains of six tones in succession as in the free-field stimulation, during evoked-response testing the same 1 s tones were presented individually, in pseudo-random order, with a stimulus-onset asynchrony (SOA) of 5 s. This resulted in the same mean presentation rate (12 tones/minute) or duty cycle (20%) as in the free-field stimulation.
The target pips had a duration of 25 ms and a frequency of 3000 Hz. They were delivered in pseudo-random order and intervals in one sixth of the inter-tone intervals at 95 dB (A-weighting), yielding a mean rate of two targets per minute. Subjects were instructed to keep their eyes closed and to press a response button each time they heard a target pip. The steady-state stimulus train consisted of 980 Hz tone pips (5 ms cosine squared envelope) at 40 dB nHL with a repetition rate of one eighth the EEG sampling rate. Its purpose was to drive a steady-state response (SSR) at the 39 Hz stimulus rate (Galambos et al., 1981), to test whether changes during exposure could be detected in the SSR.
Recording
EEG was collected from a scalp electrode
located midway between Cz and Fz of the International 10-20 System
referenced to the left earlobe.
Horizontal and lateral eye movements were also recorded.
EEG recording was continuous, with
a sampling rate of 312.5 Hz per channel
using a 12-bit analog-to-digital converter
and an analog pass band of 0.1-100 Hz.
[1]
Analysis
For each run, EEG epochs of 5.94 seconds beginning 1.23 seconds
before the onset of those tones not followed by a target pip
were selected from the continuous EEG records.
After responses containing significant eye movement or muscle artifacts
were rejected for averaging,
ERSP to each of the six tone frequencies were computed.
To compute the ERSPs,
each response epoch
was submitted to short-time Fourier analysis
using Fast Fourier transforms (FFTs).
Each epoch of 1856 points comprised 26 Hanning-windowed,
256-point data windows with 75% overlap.
Output consisted of 129 amplitude estimates with a frequency bin width
of 1.22 Hz.
To reduce random error, the 26 spectral estimates for each epoch were smoothed
using a 3-window moving-average,
resulting 24 smoothed estimate with a total spectral window length of 1.23 s.
Each moving-average spectral estimate was then normalized
by dividing by the first smoothed spectral estimate,
representing the 1.23 s pre-stimulus baseline period.
Finally, a mean subject ERSP was computed for each run
by averaging dB (20log10) transforms
of the individual-epoch ERSPs for the six tones.
The smoothing, normalization, and log averaging produced
ERSP estimates of mean dB increase or decrease in spectral
EEG amplitude timelocked to the stimulus.
[2]
Similar ERSP transforms of target stimulus-locked response epochs were also
computed and averaged.
To avoid overlap with ERSP activity to succeeding tones,
only those targets presented at least 2.8 s
before the beginning of the next tone were used in the analysis.
Target response epochs began 1.23 before
and ended 2.8 s after target onset.
Targets not followed by a button press, and target epochs containing
gross muscle artifacts were rejected for averaging.
Behavioral Thresholds
Results of audiometric testing
are more fully reported elsewhere (Krebs et al., 1992).
No significant (>=5 dB) changes in threshold were found
during or after exposure
for any subject at any frequency tested.
Nor was there any consistent change in thresholds with increasing time
of exposure, relative to pre-exposure measures.
Thus the intensity and duty-cycle of the sound the subjects were exposed to
were not sufficient to produce temporary threshold shifts (TTS).
Tone-induced ERSPs
Figure 2 shows the grand mean ERSP planes
averaged across the 28 subjects for each of the four days.
These have three main features:
1)
An augmentation in the 2-8 Hz range, peaking near 4 Hz soon after tone onset,
followed by a longer-lasting diminution in the same frequency range.
2)
A ridge of 10-40 Hz augmented activity,
with peak frequencies near 11 and 19 Hz,
emerging during the latter portion of tone,
and reaching a maximum shortly after tone offset.
3)
A suppression of EEG amplitude at the 39 Hz SSR driving frequency
during tone presentation.
>
Fig. 2:
ERSPs induced by 1 s task-irrelevant tones on Days 1-4;
grand means of responses from 28 subjects for all six tone frequencies
(720-1120 Hz). In each plot, the left-right axis is EEG frequency;
the front-back axis is time relative to tone onset (at time 0);
the vertical axis is EEG amplitude ratio plotted in (20log10) dB.
(a, b) The mean ERSP for Day 1 is closely replicated on Day 2.
(c)) Note the smaller 11-19 Hz enhancement during free-field exposure
on Day 3, and its partial recovery in (d) after exposure on Day 4.
Note that the 11-19 Hz ridge of augmented activity
appears to be smaller during free-field exposure (Day 3) than
on the other days. The actual difference between the ERSP planes
for Days 2 and 3 is shown in Figure 3a.
>
Figure 3. (a) Difference between the tone-induced ERSPs
for Day 2 (prior to exposure) and Day 3 (during exposure).
Note the relative suppression of enhancement near 11 Hz, and the
more sustained suppression of enhancement near 19 Hz on Day 3.
(b) Plot of F-statistics, showing
points of significant difference
between tone-induced ERSPs for Days 2 and 3, computed by ANOVA
from responses of 28 subjects
for each point in the time/frequency ERSP plane.
Baseline: (F(1,27)=13.5, p=.001).
To evaluate the consistency of ERSP differences
across subjects, F-statistics were computed separately
for each time/frequency point in the ERSP planes
by analysis of variance on ERSP data for each pair of Days.
To reduce the likelihood of false positive findings
due to multiple comparisons, a minimum significance level of p<.001 was used.
For the comparison between Pre-Exposure measures (Day 2 versus Day 1),
there were no points of significant difference in the ERSP plane.
By contrast, Figure 3b plots a map of F-statistics
on ERSP amplitude differences between Days 2 and 3.
In this figure
only significant F-values are shown (F(1,27)>13.5, p<.001).
Significant ERSP differences appear
near 8 Hz, soon after tone onset,
and near 11 and 19 Hz near tone offset.
Note that although the F-statistic map
has sharply defined time-frequency features,
the ERSP difference plane
(Fig. 3) shows that sizable differences in the grand means occur
over a broad range of frequencies between 8 and 25 Hz,
and that the most significant frequencies
are at the two angular peak frequencies
(near 11 and 19 Hz) visible in each of the four ERSP planes (Fig. 2).
Note also (Fig. 2d)
that recovery in the ERSPs recorded during the day after exposure (Day 4)
appears to be incomplete.
F-statistic plots of ERSP differences between Days 2 and 4 (not shown)
imply continued reductions in spectral reactivity
on the day after exposure
at about 19 Hz near tone offset,
and at about 8 Hz near tone onset.
The 8 Hz difference does not occur at a peak of either of
the ERSP planes.
Linear regression of ERSP amplitudes on time of testing
revealed no significant trends over time at any point in the ERSP plane,
either during the day of exposure (Day 3) or on the day of recovery (Day 4).
Since time-of-exposure on Day 3 (and time post-exposure on Day 4) varied
across subjects from 3 to 11 hours,
this result suggests that the psychophysiological
effects of exposure were not incremental over this time frame,
and that the effects of exposure on the ERSP
may have lasted longer than 11 hours after the end of exposure.
Target-induced ERSPs
Figure 4 shows grand mean ERSPs to detected targets.
The structure of the target-induced ERSP planes is similar to that
of the tone-induced ERSPs, although
the target responses are larger
(note the different scales used in Figs. 2 and 4).
Fig. 4:
ERSPs induced by occasional 25 ms target tonepips (at time 0)
on Days 1-4; grand means of responses from 28 subjects.
Only detected targets not followed by any stimulus for 2.6 s were
averaged.
Note the larger amplitude scale than in Fig. 2.
No tonepip response differences were found
between target ERSPs prior to and during exposure.
Both ERSP responses, to the 1 s tones and the 25 ms tonepips,
include two prominent regions of augmented activity,
a peak enhancement at 2-8 Hz following stimulus onset,
and a ridge of enhanced amplitude between 10 to 25 Hz,
peaking a second later.
In all four responses, a band of diminished amplitudes
between 20 and 30 Hz appears, peaking at about 500 ms,
with apparent minima near 24 and 29 Hz.
Target-induced ERSPs show little evidence of suppression of 39 Hz EEG
following the target tonepips.
Note in Fig. 4 that, contrary to the tone-induced ERSP results,
there is no apparent difference
between the grand mean target-induced ERSPs on the day of exposure (Day 3)
and the other days.
Statistical analyses confirmed
that the (Day 3 - Day 2) difference
was not significant (p<.001) at any point in the ERSP planes.
Significance of the ERSP
Interest in event-related EEG dynamics
other than those visible in the ERP
has been heightened in recent years by increased appreciation of
the extent of rhythmic activity induced in sensory cortices
by stimulus events.
These rhythms may be highly phase-coherent
across selected spatially adjacent or non-adjacent cortical areas,
but remain phase-incoherent across trials
and thus may not appear in the averaged evoked response
(Gray and Singer, 1989; Llinas, 1989; Murthy and Fetz, 1992).
Freeman and associates (Freeman, 1975; Freeman and Skarda, 1985),
as well as others (Basar, 1980; Grossberg, 1980),
have emphasized that
in the process of perception,
laminar brain structures, possibly including auditory cortex,
may function as low-dimensional dynamic systems
which match patterns of incoming stimulus-evoked activity against
dynamically created and maintained impressional templates stored
as a pattern of cell connectivities.
In these networks, spatially coherent, narrow-band event-related oscillations
may be a necessary concomitant of perception and object recognition,
but still may not appear in time-domain average ERPs
if their between-trials phase coherence is low.
The ERSP results obtained in this experiment
indicate that the total brain electrophysiological response
to auditory stimuli
recorded from the human scalp
is not fully revealed in the averaged ERP.
In addition to the well-known time- and phase-locked auditory ERP components,
auditory stimuli also produce
time-locked but phase-incoherent changes in the EEG spectrum,
which are revealed in the ERSP.
In this experiment, wide-band auditory ERSPs
induced by 1 s tones and by 25 ms target tonepips
shared several common features --
numerically small (1-6 dB) but replicable
enhancements of EEG amplitude timelocked to stimulus onset,
particularly in the frequency ranges 2-8 and 10-40 Hz.
In two frequency regions (8 and 11-19 Hz),
tone-induced ERSP features were significantly depressed
following sustained free-field sound exposure.
Some further discussion of these results follows,
organized by EEG band.
Delta and Theta Bands
ERSP effects below 2 Hz
were not computed in these analyses,
since accurate observation of low frequency ERSP activity
requires using a longer transform.
Near-DC ERSP activity, if demonstrated, might arise in part from
variability in slow wave components of the ERP.
Both the tone and target-induced ERSPs have a peak
in the 2-8 Hz region near tone onset.
This is the same time and frequency as the major peak in the spectrum of the
auditory ERP (Makeig, 1990; Pantev et al., 1991).
Thus the early theta band ERSP peak must include the
energy in the phase-locked ERP,
possibly enhanced by contributions from trial-to-trial ERP variance.
However, distinguishing between ERP variance
and EEG amplitude modulation models
requires an exact model of ERP variance,
since otherwise either explanation (ERP variance or EEG modulation)
can be used to fit any between-trial differences.
Alpha Band
Although an alpha rhythm generated in the auditory cortex has recently
been discovered
using biomagnetic sensors,
and has been shown to be suppressed during mental tasks,
this rhythm is not normally seen in electrical
scalp recordings, since the standard electrode montage is not
optimum for recording it
(Kaufman et at., 1989).
In this experiment, therefore,
the lack of event-related EEG suppression near 10 Hz
is not unexpected.
Pfurtscheller and Klimesch (1991) also show
an example of apparent augmentation at 10-12 Hz
after offset of a visual stimulus, an effect they explain as
relating to "disinhibition" in cortical fields.
It is unclear whether the circa 11 Hz peak in the "ridge"
feature of the ERSPs from these experiments
represent enhancement of the subject's peak alpha frequency
in the ongoing EEG, or whether it represents a separate, somewhat
higher frequency phenomenon.
Beta Band
Beta band EEG has long been assumed to represent engagement of
cortical processing (Giannitrapani, 1971),
though specific models of the role and
nature of this engagement have not been advanced.
During exposure, suppression of the beta-band augmentation was not
uniform; the peak frequencies in the pre-exposure ERSPs (near 11 and 19 Hz)
were also those most significantly reduced during exposure.
Note that the prominent 11-19 Hz ridge
of augmented activity occurring
in these ERSP experiments was induced during performance of a simple
task requiring subjects' attention to the stimuli.
The ERSP enhancement was larger
following targets than following tones,
even though the tones had a longer duration and
were perceived by subjects as being at least as intense as the tonepips.
The 11-19 Hz augmentation might, therefore, be found to vary as
a function of task and attention,
an issue which will be tested in future experiments.
If attention or other cognitive variables are shown to
affect the beta-band ERSP,
this might suggest that
human beta-band activity has a function akin to that
proposed for gamma-band processes in animals (Gray et al., 1992).
The 20-30 Hz basin of diminished amplitude
in the target pip responses near 500 ms
is in the range of the button press to these targets.
This feature might therefore reveal
movement-related suppression of coherent prestimulus
beta band activity in some or all subjects (Bouyer et al, 1980).
Gamma Band
In the human auditory cortex,
the onset of tone or click stimuli generates
a burst of 30-35 Hz of gamma band activity.
The SSR represents primarily the overlapping sum of the phase-locked
portion of these
gamma-band oscillations evoked in the auditory cortex
(Makeig, 1985, 1990; Pantev et al., 1991;
Makela and Hari, 1987).
The most prominent and consistent component in the tone-induced ERSPs (Fig. 2)
is a suppression of EEG amplitude at the steady-state stimulus rate (39 Hz)
during tone presentation.
This no doubt represents the expected suppression of
the steady-state response (SSR)
by the tone
(Makeig and Galambos, 1992a, 1992b).
Note, however, that the 39 Hz ERSP component
did not differ between Days 2 and 3 (Figs. 2b, 2c)
and thus was not affected by the free-field exposure.
This suggests that in these experiments, 3-11 hours of free-field exposure
did not alter the amplitude of gamma-band evoked responses
in the auditory cortex.
Gray and Singer (1989) have reported that a sizable subset of cells in
the visual cortices of cats respond
to moving visual stimuli
with gamma band bursts of firing with a median frequency near 50 Hz.
Murthy and Fetz (1992) have recently reported similar phase-coherent bursts
near 30 Hz in the sensorimotor cortex of
monkeys performing a difficult tactile task.
These results parallel the phenomena long observed in the olfactory cortex,
in which during inhalation of recognized odors,
40-90 Hz bursts appear (Freeman, 1975).
All these gamma band induced rhythmicities
have been theorized to provide a mechanism for rapid synchronization of
brain activation of networks of cells responsive to
different features of an object or event (so-called stimulus binding)
(Crick and Koch, 1990).
Freeman and Skarda (1985) and others (Bressler, 1990) speculate
that the occurrence of gamma-band bursts is related to the meaning
of a complex stimulus, and relates specifically to the process of forming
higher-order percepts.
In Fig. 4, it appears that the ridge of augmented 11-19 Hz activity near 1.5 s
is accompanied by a simultaneous, though smaller, increase in activity
in the 20-40 Hz range,
and preceded by an amplitude trough peaking near 500 ms at 24 and 29 Hz,
near the time of the button presses.
Why do the target-induced ERSPs not reveal evidence of
augmented higher frequency activity near stimulus onset (Makeig, 1990)?
It is likely that the time resolution of the spectral transform used
was not sufficient to reveal the existence of bursts
induced by target pips but with a much shorter duration than the
820 ms FFT windows used in the analysis.
Future ERSP analysis might study this question using
finer or variable time resolution methods
(Przybyszewski, 1991).
The Auditory ERSP
This study introduces a wideband measure of event-related
brain dynamics, the event-related spectral perturbation (ERSP),
and demonstrates
a new class of effects of exposure to sound on the brain.
ERSPs induced by two very different auditory stimuli
appear to have several time-frequency features in common,
possibly reflecting the activity of common brain response systems.
The similarities between the tone- and target-induced ERSPs
in this experiment
suggest that similar ERSP features may be induced by a wide variety
of auditory stimuli.
Note that the 1.0-1.5 s latency of the beta-band augmentation
does not seem to depend on the duration of the stimulus,
and, therefore, in the tone-induced ERSPs it is not a response to tone offset.
The early 2-8 Hz ERSP peak may represent, in part,
spectral power in the ERP itself, and, in part,
trial-to-trial variability in the phase-locked ERP.
The later beta-band component
is less likely to arise from response variability,
since there are no corresponding peaks in the ERP spectrum in this time period.
This component, therefore,
may represent new or enhanced beta- and gamma-band oscillatory activity
peaking a second or more after stimulus onset,
i.e. later than the principal components of the ERP.
As the physiological origins and psychological concomitants
of the ERSP components are not yet known,
speculations about the meaning of differences in tone-induced ERSPs
before, during, and after free-field exposure must be tentative.
These differences cannot be due to habituation in the
inner ear or peripheral auditory pathways,
since these would most probably have been accompanied
by changes in behavioral thresholds.
Intensive audiometric testing performed during this experiment
failed to reveal any effects of free-field exposure on auditory thresholds.
This suggests that the exposure produced
no lasting changes in the peripheral auditory system,
and no change in the information delivered
to those central pathways serving signal detection.
Increasing time of exposure might be expected to produce increasing
effects on auditory spectral reactivity.
However, no significant linear trends in ERSPs were found
for any Day of testing.
This suggests that 3 hours of exposure
(the minimum prior to testing for any subject)
was sufficient to produce the ERSP changes observed on Day 3.
That some ERSP changes remained on Day 4 (3-11 hours after the end of
exposure) also implies that reactive stress or arousal
associated with the onset of exposure
was not the cause of the changes observed in the tone-induced ERSPs on Day 3,
since stress might be expected to diminish on Day 3,
as subjects became habituated to their new environment,
and to ebb rapidly on Day 4 after exposure ended.
ERP / ERSP
The ERSP measures event-related
perturbations in EEG amplitudes,
but, as computed here,
necessarily includes
the spectral energy of the ERP itself.
[3]
To remove this confound,
EEG epochs timelocked to the stimulus
might first be averaged to form an ERP.
This mean ERP could then be subtracted from
the response epoch prior to analysis.
However, removing the mean ERP from each epoch before spectral analysis
would involve an implicit assumption
that event-related brain dynamics can be modeled as
the sum of a stable ERP
and a reliable pattern of EEG amplitude modulation.
Such a model would not take into account
EEG phase-dependent interactions (Jansen and Brandt, 1991),
and further, the actual effect of subtracting the mean ERP
from each epoch prior to the ERSP transform would be relatively small,
particularly at frequencies above 10 Hz
where auditory ERP amplitudes are 15 dB or more
below mean EEG amplitudes.
While the ERSP reveals new and potentially important information
about event-related brain dynamics, it cannot reveal
interactions between the ERP amplitude, latency variability,
and EEG spectral modulation.
[4]
Removing a mean ERP from each ERSP epoch prior to analysis
would not eliminate the effects on the ERSP of
trial-to-trial variability in the ERP itself.
For example, a skewed distribution of between-trial ERP variabilities
could also create the appearance of augmented EEG amplitude
in an ERP-corrected ERSP.
These complexities cannot be resolved by ERSP analysis alone
without detailed models for ERP variance and EEG modulation.
Mixed models in which the phase coherence
of evoked-response components is neither one (phase locked ERP)
nor zero (independent EEG) are also possible.
In such models the relationship between pre- and post-event EEG phase
could also be perturbed by an experimental event
(Jansen and Brandt, 1991; Achimowicz, 1992), and this could produce
additional ERSP effects.
Further Questions
The sensitivity of the tone-induced ERSP to prior stimulation,
in the absence of peripheral effects,
suggests that ERSP measures might prove sensitive to
the effects of previous experience on brain dynamics.
A number of further questions
are raised by the ERSP results of this study:
1. Prior to this experiment,
data from a group of ten subjects were collected using
an exposure and intensity of 83 dB (A-weighting).
The ERSPs for this group were generally similar to those reported here,
as were the differences between the ERSPs before and during exposure.
However, for the 83 dB group these changes were smaller and less significant,
possibly because of the smaller number of subjects.
Little else is known about the effects of stimulus intensity on the ERSP.
2. The tone frequencies used for evoked-response testing
were the same as those used in free-field exposure.
It is not known whether ERSPs to tones of different frequencies
would also be suppressed during exposure.
3. Throughout these experiments subjects
were attending the tones in anticipation of responding
to the occasional targets.
The ERSPs induced by target pips
were larger than those induced by tones,
possibly on account of their comparative probability or task relevance.
Effects of alertness, task relevance, and attention
on ERSP form and amplitude remain to be explored (Makeig and Inlow, 1992).
4. Averaging over all subjects and trials
yields information about only
the most global event-related response patterns--
between-subject and between-trials differences are ignored.
Gray et al. (1992) have recently
shown that spatial coherences among event-related gamma band oscillations
in the visual cortex of cats are complex transitory dynamic phenomena.
It is probable that single-trial ERSPs reported here also
represent transient and somewhat variable dynamic events.
Cluster analysis of subjects and trials,
followed by discrete or moving subaverages,
might prove useful for
better understanding the brain processes measured by ERSPs.
1.
A wideband, normalized measure of event-related spectral reactivity, the
event-related spectral perturbation or ERSP,
can be used to measure aspects of event-related brain dynamics
not found in the ERP.
2.
Auditory ERSPs
had similar forms and latencies for two very different inducing stimuli --
1 s, low-frequency tones, and 25 ms, 3 kHz tone pips.
A prominent feature of both ERSPs was a prominent, relatively late (1.0-1.5s)
ridge of augmented 11-19 Hz EEG amplitudes
which was larger in response to the target tonepips.
3.
Some features of the auditory ERSP appear to be sensitive to
previous sound exposure.
In this experiment, some features of the tone-induced ERSPs
were significantly suppressed
following prolonged exposure to moderately loud pure tones,
and appeared to not recover fully
on the day following exposure.
The physiological significance of the ERSP features revealed in these
experiments is not yet known,
and many experiments may be required to determine their properties.
Auditory, visual and somatosensory stimulus-induced ERSPs
may be expected to differ in form, time course, and scalp distribution.
Analysis of single response epochs,
multi-channel source localization studies, and multi-modality
experiments would also be of interest.
Results of cognitive experiments,
including experiments involving omitted stimuli,
could add to our understanding of the relation of EEG activity to cognition.
It is likely that further research using wide band ERSP measures can
yield new information on brain dynamics of perception
and cognition.
ACKNOWLEDGEMENTS
The author wishes to acknowledge the assistance of
Steven Linnville, Anne Makeig and Candee Corwin in collecting the data,
of Mark Inlow in discussing the results,
and of Robert Galambos and Jerszy Achimowicz
in commenting on earlier versions of this report.
Attias,
J., and Pratt, H. "Auditory-evoked potential correlates
of susceptibility to noise-induced hearing loss,"
Audiology 1985,24, 149-156.
Basar,
A.E.
EEG Brain Dynamics, (Elsevier, Amsterdam), 1980.
Bouyer,
J.J. Montaron, M.F., Rougeul-Buser, A., and Buser, P.,
"A thalamo-cortical rhythmic system accompanying high vigilance
levels in the cat,"
in Rhythmic EEG Activities and Cortical Functioning, pp.
edited by G. Pfurtscheller
(Elsevier/North_holland Biomedical Press, Amsterdam),
pp. 63-77, 1980.
Bressler,
S.L. "The gamma wave: a cortical information
carrier?," Trends in Neuroscience 1990, 13(5), 161-162.
Clark,
W.W. "Recent studies of temporary threshold shift (TTS) and
permanent threshold shift (PTS) in animals,"
J. Acoust. Soc. Amer. 1991, 90,155-163.
Coyne,
J.T., ed.
Investigation of shipboard habitability during low frequency active
(LFA) sonar operations.
Technical Report, Naval Health Research Center, San Diego, CA, 1992.
Crick,
F. and Koch, C. "Towards a neurobiological
theory of consciousness," Seminars in the Neurosciences 1990, 2,263-275.
Dejoy,
D.M., "The nonauditory effects of noise: review
and perspectives for research,"
J. Auditory Res. 1984, 24,123-150.
Freeman,
W. Mass Action in the Nervous System,
(Academic Press, New York), 1975.
Freeman,
W.J. and Skarda, C.A. "Spatial EEG patterns,
non-linear dynamics and perception: the neo-Sheringtonian view,"
Brain Res. Rev. 1985, 10, 147-175.
Galambos,
R., Makeig, S., and Talmachoff, P. "A 40 Hz
auditory potential recorded from the human scalp,"
Proc. Nat. Acad. Sci. USA 1981, 78(4), 2643-2647.
Galambos,
R., and Makeig, S.,
"Studies of central masking I. Effects of noise
on the steady-state response,"
J Acoust Soc Amer, 1992a.
Galambos,
R., and Makeig, S.,
"Studies of central masking II. Use of tonal probes
and a masking level difference experiment,"
J Acoust Soc Amer, 1992b.
Giannitrapani,
A.D.
"Scanning mechanisms and the EEG,"
Electroenceph. clin. Neurophysiol.
1971, 30,139-146.
Gray,
C.M. and Singer, W. "Stimulus-specific neuronal
oscillations in orientation columns of cat visual cortex,"
Proc. Nat. Acad. Sci. USA, 1989, 86,1698-1702.
Gray,
C.M., Engel, A.K., Konig, P., and Singer, W.
"Synchronization of oscillatory neuronal responses in cat striate cortex:
Temporal properties,"
Visual Neuroscience 1992, 8,337-347.
Grossberg,
S.
"How does a brain build a cognitive code?,"
Psychol. Rev.
1980, 87,1-51.
Jansen,
G.
"Physiological effects of noise,"
in Handbook of Acoustical Measurements and Noise Control, 3rd ed.,
edited by C. Harris,
(McGraw Hill, New York),
pp. 25.1-25.19, 1991.
Jansen,
B.H. and Brandt, M.E.
"The effect of the phase of prestimulus alpha activity
on the averaged visual evoked response,"
Electroenceph. clin. Neurophysiol.
1991, 80, 241-250.
Jones,
D. and Broadbent, D. "Human performance and noise."
in Handbook of Acoustical Measurements and Noise Control, 3rd ed.,
edited by C. Harris,
(McGraw Hill, New York),
pp. 18.1-18.19, 1991.
Kaufman
L., M. Glanzer, Y. Cycowicz, and S. Williamson
"Visualizing and rhyming cause differences in alpha suppression,"
in Advances in Biomagnetism.
edited by S. Williamson, M. Hoke, G. Stroink, and M. Kotani, eds.
(Plenum Press, New York), pp. 241-24, 1989.
Krebs,
D., Galambos, R., and Gales, R.S.
The effect of LFA sonar transmissions upon human hearing.
Technical Report, Naval Health Research Center, San Diego, CA, 1992.
Llinas,
R.R.
"The intrinsic electrophysiological properties
of mammalian neurons: insights into central nervous system function,"
Science 1989, 242, 1654-1664.
Makeig,
S.
Studies in Musical Psychobiology
University Microfilms, Ann Arbor, MI, 1985.
Makeig,
S. and Galambos, R.
"The CERP: Event-related
perturbations in steady-state responses,"
in Brain Dynamics: Progress and Perspectives,
edited by E. Basar and T. Bullock
(Springer, Berlin), pp. 375-400, 1989.
Makeig,
S.
"A dramatic increase in the auditory middle latency
response at very slow rates,"
in Psychophysiological Brain Research,
edited by C. Brunia, G. Mulder and M.N. Verbaten,
(Tilburg University Press, Tilburg, The Netherlands),
Vol.2, pp. 60-64, 1990.
Makeig,
S. and Inlow, M.,
"Lapses in alertness: coherence of
fluctuations in performance and EEG spectrum,"
Electroenceph. clin. Neurophysiol, in press.
Makela,
J.P. and Hari, R.
"Evidence for cortical origin of the 40 Hz auditory evoked response in man,"
Electroenceph. clin. Neurophysiol.1987, 66,539-546.
Murthy,
V.S. and Fetz, E.E.
"Coherent 25-35 Hz
oscillations in the sensorimotor cortex of awake behaving monkeys,"
Proc. Nat. Acad. Sci. USA 1992, 89, 5670-5674.
Melnick,
W. "Hearing loss from noise exposure,"
in Handbook of Acoustical Measurements and Noise Control, 3rd ed.,
edited by C. Harris,
(McGraw Hill, New York),
pp. 18.1-18.19, 1991.
Pantev,
C., Makeig, S., Hoke, M., Galambos, R., Hampson, S. and
Gallen, C.
"Human auditory evoked gamma band magnetic fields,"
Proc. Nat. Acad. Sci. USA 1991, 88,8996-9000.
Pfurtscheller,
G.
"Graphical display and statistical
evaluation of event-related desynchronization (ERD),"
Electroenceph. clin. Neurophysiol. 1977, 43, 757-750.
Pfurtscheller,
G. and Klimesch, W.
"Event-related
desynchronization during motor behavior and visual information processing."
in Event-Related Brain Research (EEG Suppl. 42)
edited by C.H.M. Brunia, G. Mulder and M.N. Verbaten,
(Elsevier Science Publishers, Amsterdam), pp. 58-65, 1991.
Pfurtscheller,
G.
"Event-related synchronization (ERS): an electrophysiological correlate
of cortical areas at rest,"
Electroenceph. clin. Neurophysiol. 1992, 83, 62-69.
Przybyszewski,
A.W.
"An analysis of the oscillatory
patterns in the central nervous system with the wavelet method,"
J. Neurosci. Methods 1991, 38, 247-257.
Van Dijk,
J. G., Caekebeke, J., Jennekens-Schinkel, A., and Zwinderman, A.
"Background EEG reactivity in auditory event-related potentials,"
Electroenceph. clin. Neurophysiol. 1992, 83, 44-51.
1.
Subsequent to data collection, it was found that because of a
minor computer malfunction, stimuli and EEG recording
had not been precisely synchronized.
Using steady-state response (SSR) phase results, it was determined
that stimulus presentation lagged behind EEG data collection
at a rate of one frame in 7728.
Using this information, moments of stimulus onset were corrected post hoc.
Because ERSP analysis trades time resolution for frequency resolution,
remaining uncertainty in stimulus timing (estimated from control runs
as less than 25 ms (root mean squared))
could affect the ERSP results very little.
However, ERP measures depend on more precise response phase synchronization,
and therefore are not reported here.
(Return)
2.
Amplitude/baseline amplitude normalization and mean log averaging provide one of
several possible stochastic estimates of mean tendency in the distribution
of response epochs. Median, arithmetic mean, and root mean square averaging
might also be used to give somewhat different estimates.
Averaging of event-related spectra can also be carried out without normalizing,
producing an event-related spectrum transform.
However, this can be dominated
by noisy large-amplitude epochs in which the tendency for proportionate
amplitude increase or decrease is obscured or absent.
(Return)
3.
This confound between EEG and ERP spectral characteristics
is also present in narrow-band ERDs.
(Return)
4.
In general, vestiges of phenomena revealed by ERSP analysis can remain
in ERPs as time-dependent changes
in the spectral content of the background EEG "noise"
remaining in the ERP average.
However, as ERP analysis is designed to remove phase-incoherent
activity occurring in the response period,
the vestiges of the ERSP remaining in the ERP are normally small,
and in general ERSP features cannot be expected
to be recaptured efficiently from the ERP by
post hoc spectral analysis.
(Return)
RESULTS
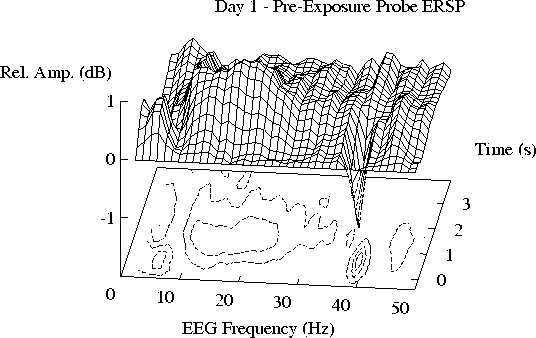
Fig. 2aSorry, Figure 2b not yet available by www...
Fig. 2b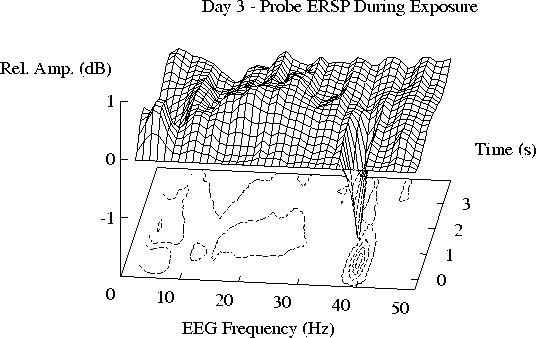
Fig. 2c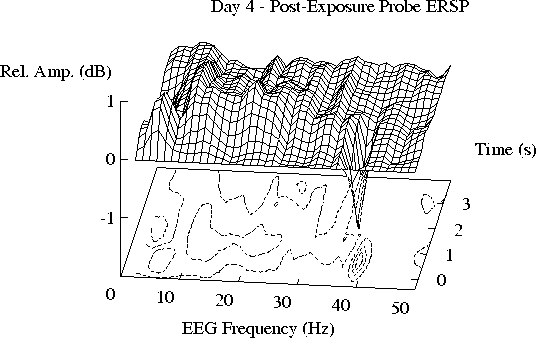
Fig. 2d
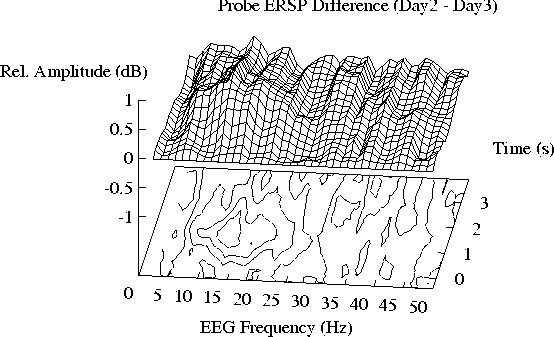
Fig. 3aSorry, Figure 3b not yet available by www...
Fig. 3b
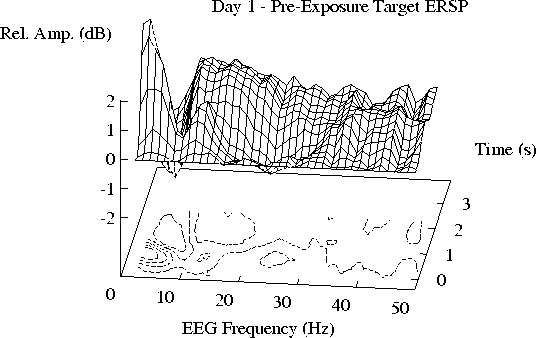
Fig. 4a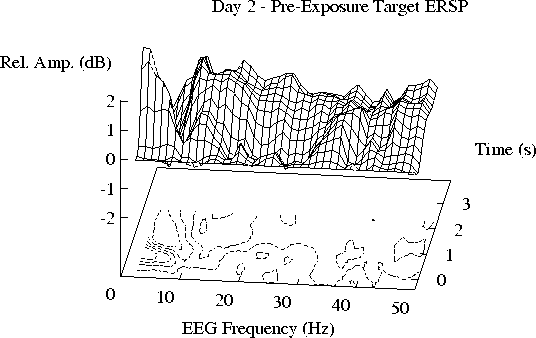
Fig. 4b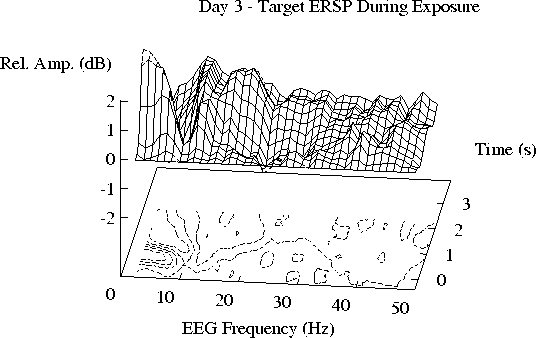
Fig. 4c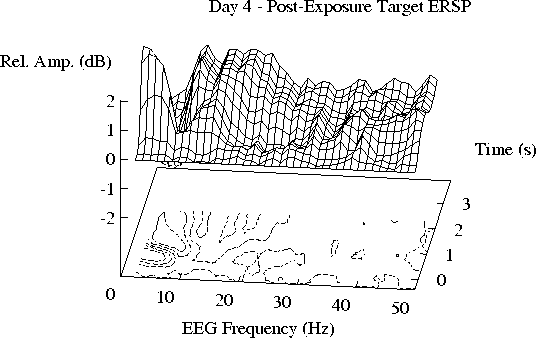
Fig. 4d
DISCUSSION
GENERAL DISCUSSION
CONCLUSIONS
REFERENCES
Achimowicz,
J., "On the deterministic brain states revealed by VEP
classification in phase domain,"
in Mathematical Approaches to Brain Functioning Diagnostics,
edited by I. Dvorak and A. Holden,
(Manchester University Press, Manchester),
pp. 209-230, 1992.
FOOTNOTES